Медицинский эксперт статьи

Новые публикации
Гипофиз
Последняя редакция: 23.04.2024

Весь контент Web2Health проверяется медицинскими экспертами, чтобы обеспечить максимально возможную точность и соответствие фактам.
У нас есть строгие правила по выбору источников информации и мы ссылаемся только на авторитетные сайты, академические исследовательские институты и, по возможности, доказанные медицинские исследования. Обратите внимание, что цифры в скобках ([1], [2] и т. д.) являются интерактивными ссылками на такие исследования.
Если вы считаете, что какой-либо из наших материалов является неточным, устаревшим или иным образом сомнительным, выберите его и нажмите Ctrl + Enter.
Гипофиз (hypophysis, s.glandula pituitaria) находится в гипофизарной ямке турецкого седла клиновидной кости и отделен от полости черепа отростком твердой оболочки головного мозга, образующим диафрагму седла. Через отверстие в этой диафрагме гипофиз соединен с воронкой гипоталамуса промежуточного мозга. Поперечный размер гипофиза равен 10-17 мм, переднезадний - 5-15 мм, вертикальный - 5-10 мм. Масса гипофиза у мужчин равна примерно 0,5 г, у женщин - 0,6 г. Снаружи гипофиз покрыт капсулой.
В соответствии с развитием гипофиза из двух разных зачатков в органе различают две доли - переднюю и заднюю. Аденогипофиз, или передняя доля (adenohypophysis, s.lobus anterior), более крупная, составляет 70-80 % от всей массы гипофиза. Она более плотная, чем задняя доля. В передней доле выделяют дистальную часть (pars distalis), которая занимает переднюю часть гипофизарной ямки, промежуточную часть (pars intermedia), расположенную на границе с задней долей, и бугорную часть (pars tuberalis), уходящую вверх и соединяющуюся с воронкой гипоталамуса. В связи с обилием кровеносных сосудов передняя доля имеет бледно-желтый, с красноватым оттенком цвет. Паренхима передней доли гипофиза представлена несколькими типами железистых клеток, между тяжами которых располагаются синусоидальные кровеносные капилляры. Половина (50 %) клеток аденогипофиза являются хромафильными аденоцитами, имеющими в своей цитоплазме мелкозернистые гранулы, хорошо окрашивающиеся солями хрома. Это ацидофильные аденоциты (40 % от всех клеток аденогипофиза) и базофильные аденоциты {10 %). В число базофильных аденоцитов входят гонадотропные, кортикотропные и тиреотропные эндокриноциты. Хромофобные аденоциты мелкие, они имеют крупное ядро и небольшое количество цитоплазмы. Эти клетки считаются предшественниками хромофильных аденоцитов. Другие 50 % клеток аденогипофиза являются хромофобными аденоцитами.
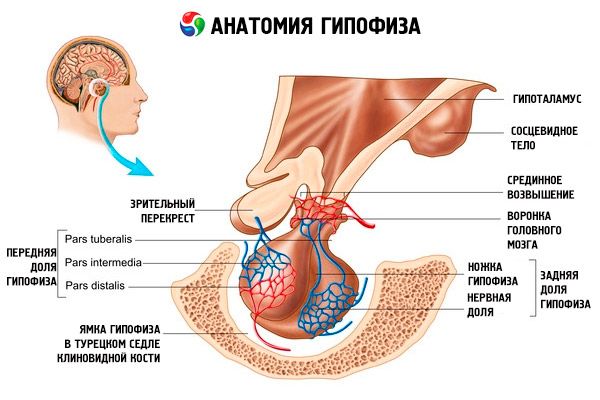
Нейрогипофиз, или задняя доля (neurohypophysis, s.lobus posterior), состоит из нервной доли (lobus nervosus), которая находится в задней части гипофизарной ямки, и воронки (infundibulum), расположенной позади бугорной части аденогипофиза. Задняя доля гипофиза образована нейроглиальными клетками (питуициты), нервными волокнами, идущими от нейросекреторных ядер гипоталамуса в нейрогипофиз, и нейросекреторными тельцами.
Гипофиз при помощи нервных волокон (путей) и кровеносных сосудов функционально связан с гипоталамусом промежуточного мозга, который регулирует деятельность гипофиза. Гипофиз и гипоталамус вместе с их нейроэндокринными, сосудистыми и нервными связями принято рассматривать как гипоталамо-гипофизарную систему.
Гормоны передней и задней долей гипофиза оказывают влияние на многие функции организма, в первую очередь через другие эндокринные железы. В передней доле гипофиза ацидофильные аденоциты (альфа-клетки) вырабатывают соматотропный гормон (гормон роста), принимающий участие в регуляции процессов роста и развития молодого организма. Кортикотропные эндокриноциты секретируют адренокортикотропный гормон (АКТГ), стимулирующий секрецию стероидных гормонов надпочечниками. Тиротропные эндокриноциты секретируют тиротропный гормон (ТТГ), влияющий на развитие щитовидной железы и активирующий продукцию ее гормонов. Гонадотропные гормоны: фолликулостимулирующий (ФСГ), лютеинизирующий (ЛГ) и пролактин - влияют на половое созревание организма, регулируют и стимулируют развитие фолликулов в яичнике, овуляцию, рост молочных желез и выработку молока у женщин, процесс сперматогенеза у мужчин. Эти гормоны вырабатываются базофильными аденоцитами бета-клетки). Здесь же секретируются липотропные факторы гипофиза, которые оказывают влияние на мобилизацию и утилизацию жиров в организме. В промежуточной части передней доли образуется меланоцитостимулирующий гормон, контролирующий образование пигментов - меланинов - в организме.
Нейросекреторные клетки супраоптического и паравентрикулярного ядер в гипоталамусе продуцируют вазопрессин и окситоцин. Эти гормоны транспортируются к клеткам задней доли гипофиза по аксонам, составляющим гипоталамо-гипофизарный тракт. Из задней доли гипофиза эти вещества поступают в кровь. Гормон вазопрессин оказывает сосудосуживающее и антидиуретическое действие, за что и получил также название антидиуретического гормона (АДГ). Окситоцин оказывает стимулирующее влияние на сократительную способность мускулатуры матки, усиливает выделение молока лактирующей молочной железой, тормозит развитие и функцию желтого тела, влияет на изменение тонуса гладких (неисчерченных) мышц желудочно-кишечного тракта.
Развитие гипофиза
Передняя доля гипофиза развивается из эпителия дорсальной стенки ротовой бухты в виде кольцевидного выроста (карман Ратке). Это эктодермальное выпячивание растет в сторону дна будущего III желудочка. Навстречу ему от нижней поверхности второго мозгового пузыря (будущее дно III желудочка) вырастает отросток, из которого развиваются серый бугор воронки и задняя доля гипофиза.
Сосуды и нервы гипофиза
От внутренних сонных артерий и сосудов артериального круга большого мозга к гипофизу направляются верхние и нижние гипофизарные артерии. Верхние гипофизарные артерии идут к серому ядру и воронке гипоталамуса, анастомозируют здесь друг с другом и образуют проникающие в ткань мозга капилляры - первичную гемокапиллярную сеть. Из длинных и коротких петель этой сети формируются воротные вены, которые направляют к передней доле гипофиза. В паренхиме передней доли гипофиза эти вены распадаются на широкие синусоидальные капилляры, образующие вторичную гемокапиллярную сеть. Задняя доля гипофиза кровоснабжается преимущественно за счет нижней гипофизарной артерии. Между верхними и нижними гипофизарными артериями имеются длинные артериальные анастомозы. Отток венозной крови из вторичной гемокапиллярной сети осуществляется по системе вен, впадающих в пещеристые и межпещеристые синусы твердой оболочки головного мозга.
В иннервации гипофиза участвуют симпатические волокна, проникающие в орган вместе с артериями. Постганглио-нарные симпатические нервные волокна отходят от сплетения внутренней сонной артерии. Помимо этого, в задней доле гипофиза обнаруживаются многочисленные окончания отростков нейросекреторных клеток, залегающих в ядрах гипоталамуса.
Возрастные особенности гипофиза
Средняя масса гипофиза у новорожденных достигает 0,12 г. Масса органа удваивается к 10 и утраивается к 15 годам. К 20-летнему возрасту масса гипофиза достигает максимума (530-560 мг) и в последующие возрастные периоды почти не меняется. После 60 лет наблюдается небольшое уменьшение массы этой железы внутренней секреции.
 [7], [8], [9], [10], [11], [12], [13], [14], [15]
[7], [8], [9], [10], [11], [12], [13], [14], [15]
Гормоны гипофиза
Единство нервной и гормональной регуляции в организме обеспечивается тесной анатомической и функциональной связью гипофиза и гипоталамуса. Этот комплекс определяет состояние и функционирование всей эндокринной системы.
Главная железа внутренней секреции, вырабатывающая ряд пептидных гормонов, непосредственно регулирующих функцию периферических желез, - гипофиз. Это красновато-серое образование бобовидной формы, покрытое фиброзной капсулой массой 0,5-0,6 г. Он незначительно меняется в зависимости от пола и возраста человека. Общепринятым остается деление гипофиза на две доли, различные по развитию, строению и функциям: переднюю дистальную - аденогипофиз и заднюю - нейрогипофиз. Первый составляет около 70 % от общей массы железы и условно делится на дистальную, воронковую и промежуточную части, второй - на заднюю часть, или долю, и гипофизарную ножку. Железа расположена в гипофизарной ямке турецкого седла клиновидной кости и через ножку связана с мозгом. Верхняя часть передней доли прикрыта зрительным перекрестом и зрительными трактами. Кровоснабжение гипофиза весьма обильно и осуществляется ветвями внутренней сонной артерии (верхней и нижней гипофизарными артериями), а также ветвями артериального круга большого мозга. Верхние гипофизарные артерии участвуют в кровоснабжении аденогипофиза, а нижние - нейрогипофиза, контактируя при этом с нейросекреторными окончаниями аксонов крупноклеточных ядер гипоталамуса. Первые входят в срединное возвышение гипоталамуса, где рассыпаются в капиллярную сеть (первичное капиллярное сплетение). Эти капилляры (с которыми контактируют терминали аксонов мелких нейросекреторных клеток медиобазального гипоталамуса) собираются в портальные вены, спускающиеся вдоль гипофизарной ножки в паренхиму аденогипофиза, где вновь разделяются на сеть синусоидных капилляров (вторичное капиллярное сплетение). Так, кровь, предварительно пройдя через срединное возвышение гипоталамуса, где обогащается гипоталамическими аденогипофизотропными гормонами (рилизинг-гормонами), попадает к аденогипофизу.
Отток крови, насыщенной аденогипофизарными гормонами, из многочисленных капилляров вторичного сплетения осуществляется по системе вен, которые в свою очередь впадают в венозные синусы твердой мозговой оболочки и далее в общий кровоток. Таким образом, портальная система гипофиза с нисходящим направлением тока крови от гипоталамуса является морфофункциональным компонентом сложного механизма нейрогуморального контроля тропных функций аденогипофиза.
Иннервация гипофиза осуществляется симпатическими волокнами, следующими по гипофизарным артериям. Начало им дают постганглионарные волокна, идущие через внутреннее сонное сплетение, связанное с верхними шейными узлами. Прямой иннервации аденогипофиза от гипоталамуса нет. В заднюю долю поступают нервные волокна нейросекреторных ядер гипоталамуса.
Аденогипофиз по гистологической архитектонике представляет собой весьма сложное образование. В нем различают два вида железистых клеток - хромофобные и хр.омофильные. Последние в свою очередь делятся на ацидофильные и базофильные (детальное гистологическое описание гипофиза дано в соответствующем разделе руководства). Однако следует отметить, что гормоны, продуцируемые железистыми клетками, входящими в состав паренхимы аденогипофиза, из-за многообразия последних в какой-то степени различны по своей химической природе, а тонкая структура секретизирующих клеток должна соответствовать особенностям биосинтеза каждого из них. Но иногда в аденогипофизе можно наблюдать и переходные формы железистых клеток, которые способны вырабатывать несколько гормонов. Имеются сведения о том, что разновидность железистых клеток аденогипофиза не всегда определяется генетически.
Под диафрагмой турецкого седла находится воронковая часть передней доли. Она охватывает ножку гипофиза, контактируя с серым бугром. Эта часть аденогипофиза характеризуется наличием в ней эпителиальных клеток и обильным кровоснабжением. Она также гормонально-активна.
Промежуточная (средняя) часть гипофиза состоит из нескольких слоев крупных секреторно-активных базофильных клеток.
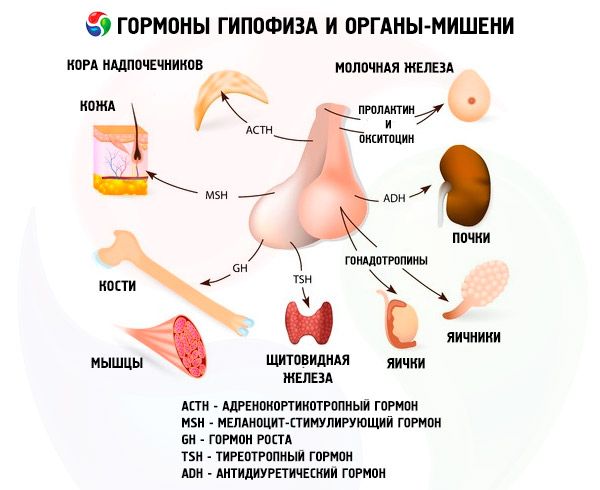
Гипофиз через свои гормоны осуществляет разнообразные функции. В его передней доле вырабатываются адренокортикотропный (АКТГ), тиреотропный (ТТГ), фолликулостимулирующий (ФСГ), лютеинизирующий (ЛГ), липотропные гормоны, а также гормон роста - соматотропный (СТО и пролактин. В промежуточной доле синтезируется меланоцитостимулирующий гормон (МСГ), а в задней накапливается вазопрессин и окситоцин.
АКТГ
Гипофизарные гормоны представляют группу белковых и пептидных гормонов и гликопротеидов. Из гормонов передней доли гипофиза наиболее изучен АКТГ. Он вырабатывается базофильными клетками. Основная его физиологическая функция - стимуляция биосинтеза и секреция стероидных гормонов корой надпочечников. АКТГ также проявляет меланоцитостимулирующую и липотропную активность. В 1953 г. он был выделен в чистом виде. В дальнейшем была установлена его химическая структура, состоящая у человека и ряда млекопитающих из 39 аминокислотных остатков. АКТГ не обладает видовой специфичностью. В настоящее время осуществлен химический синтез как самого гормона, так и различных, более активных, чем природные гормоны, фрагментов его молекулы. В структуре гормона два участка пептидной цепи, один из которых обеспечивает обнаружение и связывание АКТГ с рецептором, а другой - дает биологический эффект. С рецептором АКТГ, по-видимому, связывается за счет взаимодействия электрических зарядов гормона и рецептора. Роль биологического эффектора АКТГ выполняет фрагмент молекулы 4-10 (Мет-Глу-Гис-Фен-Арг-Три-Три).
Меланоцитостимулирующая активность АКТГ обусловлена присутствием в молекуле N-концевого участка, состоящего из 13 аминокислотных остатков и повторяющего структуру альфа-меланоцитостимулирующего гормона. Этот же участок содержит гептапептид, присутствующий в других гормонах гипофиза и обладающий некоторой адренокортикотропной, меланоцитостимулирующей и липотропной активностями.
Ключевым моментом в действии АКТГ следует считать активацию фермента протеинкиназы в цитоплазме с участием цАМФ. Фосфорилированная протеинкиназа активирует фермент эстеразу, превращающий эфиры холестерина в свободное вещество в жировых каплях. Белок, синтезированный в цитоплазме в результате фосфорилирования рибосом, стимулирует связывание свободного холестерина с цитохромом Р-450 и перенос его из липидных капель в митохондрии, где присутствуют все ферменты, обеспечивающие превращение холестерина в кортикостероиды.
[16], [17], [18], [19], [20], [21], [22]
Тиреотропный гормон
ТТГ - тиреотропин - основной регулятор развития и функционирования щитовидной железы, процессов синтеза и секреции тиреоидных гормонов. Этот сложный белок - гликопротеид - состоит из альфа- и бета-субъединиц. Структура первой субъединицы совпадает с альфа-субъединицей лютеинизирующего гормона. Более того, она в значительной степени совпадает у разных видов животных. Последовательность аминокислотных остатков в бета-субъединице ТТГ человека расшифрована и состоит из 119 аминокислотных остатков. Можно отметить, что бета-субъединицы ТТГ человека и крупного рогатого скота во многом сходны. Биологические свойства и характер биологической активности гликопротеидных гормонов определяются бета-субъединицей. Она также обеспечивает взаимодействие гормона с рецепторами в различных органах-«мишенях». Однако бета-субъединица у большинства животных проявляет специфическую активность только после соединения ее с альфа-субъединицей, выступающей в роли своеобразного активатора гормона. При этом последняя с одинаковой вероятностью индуцирует лютеинизирующую, фолликулостимулирующую и тиреотропную активности, определяемые свойствами бета-субъединицы. Обнаруженное сходство позволяет сделать заключение о возникновении этих гормонов в процессе эволюции из одного общего предшественника, бета-субъединица обусловливает и иммунологические свойства гормонов. Есть предположение, что альфа-субъединица защищает бета-субъединицу от действия протеолитических ферментов, а также облегчает транспортировку ее из гипофиза к периферическим органам-«мишеням».
Гонадотропные гормоны
Гонадотропины представлены в организме в виде ЛГ и ФСГ. Функциональное предназначение этих гормонов в целом сводится к обеспечению репродуктивных процессов у особей обоего пола. Они, как и ТТГ, являются сложными белками - гликопротеидами. ФСГ индуцирует созревание фолликулов в яичниках у самок и стимулирует сперматогенез у самцов. ЛГ вызывает у самок разрыв фолликула с образованием желтого тела и стимулирует секрецию эстрогенов и прогестерона. У самцов этот же гормон ускоряет развитие интерстициальной ткани и секрецию андрогенов. Эффекты действия гонадотропинов зависимы друг от друга и протекают синхронно.
Динамика секреции гонадотропинов у женщин меняется в ходе менструального цикла и достаточно подробно изучена. В преовуляторную (фолликулярную) фазу цикла содержание ЛГ находится на довольно низком уровне, а ФСГ - увеличено. По мере созревания фолликула секреция эстрадиола повышается, что способствует повышению продуцирования гипофизом гонадотропинов и возникновению циклов как ЛГ, так и ФСГ, т. е. половые стероиды стимулируют секрецию гонадотропинов.
В настоящее время структура ЛГ определена. Как и ТТГ, он состоит из 2 субъединиц: а и р. Структура альфа-субъединицы ЛГ у разных видов животных в значительной степени совпадает, она соответствует строению алфьа-субъединицы ТТГ.
Структура бета-субъединицы ЛГ заметно отличается от строения бета-субъединицы ТТГ, хотя имеет четыре одинаковых участка пептидной цепи, состоящих из 4-5 аминокислотных остатков. В ТТГ они локализуются в положениях 27-31, 51-54, 65-68 и 78-83. Так как бета-субъединица ЛГ и ТТГ определяет специфическую биологическую активность гормонов, то можно предположить, что гомологичные участки в структуре ЛГ и ТТГ должны обеспечивать соединение бета-субъединиц с альфа-субъединицей, а разные по структуре участки - отвечать за специфичность биологической активности гормонов.
Нативный ЛГ очень стабилен к действию протеолитических ферментов, однако бета-субъединица быстро расщепляется химотрипсином, а а-субъединица трудно гидролизуется ферментом, т. е. она выполняет защитную роль, предотвращая доступ химотрипсина к пептидным связям.
Что касается химической структуры ФСГ, то в настоящее время исследователи не получили окончательных результатов. Так же, как и ЛГ, ФСГ состоит из двух субъединиц, однако бета-субъединица ФСГ отличается от бета-субъединицы ЛГ.
Пролактин
В процессах репродукции активное участие принимает еще один гормон - пролактин (лактогенный гормон). Основные физиологические свойства пролактина у млекопитающих проявляются в виде стимуляции развития молочных желез и лактации, роста сальных желез и внутренних органов. Он способствует проявлению эффекта стероидов на вторичные половые признаки у самцов, стимулирует секреторную активность желтого тела у мышей и крыс и участвует в регуляции жирового обмена. Много внимания уделяется пролактину в последние годы как к регулятору материнского поведения, такая полифункциональность объясняется его эволюционным развитием. Он один из древних гипофизарных гормонов и обнаруживается даже у амфибий. В настоящее время полностью расшифрована структура пролактина некоторых видов млекопитающих. Однако до последнего времени ученые высказывали сомнения в существовании такого гормона у человека. Многие считали, что его функцию выполняет гормон роста. Сейчас получены убедительные доказательства наличия пролактина у человека и частично расшифрована его структура. Рецепторы пролактина активно связывают гормон роста и плацентарный лактоген, что свидетельствует о едином механизме действия трех гормонов.
Соматотропин
Еще более широким спектром действия, чем пролактин, обладает гормон роста - соматотропин. Как и пролактин, он вырабатывается ацидофильными клетками аденогипофиза. СТГ стимулирует рост скелета, активирует биосинтез белка, дает жиромобилизующий эффект, способствует увеличению размеров тела. Кроме того, он координирует обменные процессы.
Участие гормона в последних подтверждается фактом резкого увеличения его секреции гипофизом, например, при снижении содержания сахара в крови.
Химическая структура этого гормона человека в настоящее время полностью установлена - 191 аминокислотный остаток. Первичная структура его аналогична строению хорионического соматомаммотропина или плацентарного лактогена. Эти данные указывают на значительную эволюционную близость двух гормонов, хотя они проявляют различия в биологической активности.
Необходимо подчеркнуть большую видовую специфичность рассматриваемого гормона - например, СТГ животного происхождения неактивен у человека. Это объясняется как реакцией между рецепторами СТГ человека и животных, так и строением самого гормона. В настоящее время ведутся исследования по выявлению активных центров в сложной структуре СТГ, проявляющих биологическую активность. Изучаются отдельные фрагменты молекулы, проявляющие иные свойства. Например, после гидролиза СТГ человека пепсином был выделен пептид, состоящий из 14 аминокислотных остатков и соответствующий участку молекулы 31-44. Он не обладал эффектом роста, но по липотропной активности значительно превосходил нативный гормон. Гормон роста человека, в отличие от аналогичного гормона животных, обладает значительной лактогенной активностью.
В аденогипофизе синтезируется много как пептидных, так и белковых веществ, обладающих жиромобилизующим действием, а тропные гормоны гипофиза - АКТГ, СТГ, ТТГ и другие - оказывают липотропное действие. В последние годы особо выделены бета- и у-липотропные гормоны (ЛПГ). Наиболее подробно изучены биологические свойства бета-ЛПГ, который, помимо липотропной активности, оказывает также меланоцитостимулирующее, кортикотропинстимулирующее и гипокальциемическое действие, а также дает инсулиноподобный эффект.
В настоящее время расшифрована первичная структура овечьего ЛПГ (90 аминокислотных остатков), липотропных гормонов свиньи и крупного рогатого скота. Этот гормон имеет видовую специфичность, хотя структура центрального участка бета-ЛПГ у разных видов одинакова. Она определяет биологические свойства гормона. Один из фрагментов этого участка обнаруживается в структуре альфа-МСГ, бета-МСГ, АКТГ и бета-ЛПГ. Высказывается предположение, что эти гормоны в процессе эволюции возникли из одного и того же предшественника. у-ЛПГ обладает более слабой липотропной активностью, чем бета-ЛПГ.
Меланоцитостимулирующий гормон
Этот гормон, синтезирующийся в промежуточной доле гипофиза, по своей биологической функции стимулирует биосинтез кожного пигмента меланина, способствует увеличению размеров и количества пигментных клеток меланоцитов в кожных покровах земноводных. Эти качества МСГ используются при биологическом тестировании гормона. Различают два типа гормона: альфа- и бета-МСГ. Показано, что альфа-МСГ не обладает видовой специфичностью и имеет одинаковое химическое строение у всех млекопитающих. Молекула его представляет собой пептидную цепь, состоящую из 13 аминокислотных остатков. Бета-МСГ, напротив, обладает видовой специфичностью, и структура его различается у разных животных. У большинства млекопитающих молекула бета-МСГ состоит из 18 аминокислотных остатков, и только у человека она удлинена с аминного конца на четыре аминокислотных остатка. Следует отметить, что альфа-МСГ обладает некоторой адренокортикотропной активностью, и в настоящее время доказано его влияние на поведение животных и человека.
Окситоцин и вазопрессин
В задней доле гипофиза скапливаются вазопрессин и окситоцин, которые синтезируются в гипоталамусе: вазопрессин - в нейронах супраоптического ядра, а окситоцин - паравентрикуляторного. Далее они переносятся в гипофиз. Следует подчеркнуть, что в гипоталамусе вначале синтезируется предшественник гормона вазопрессина. Одновременно там же продуцируется белок-нейрофизин 1-го и 2-го типов. Первый связывает окситоцин, а второй - вазопрессин. Эти комплексы мигрируют в виде нейросекреторных гранул в цитоплазме вдоль аксона и достигают задней доли гипофиза, где нервные волокна заканчиваются в стенке сосудов и содержимое гранул поступает в кровь. Вазопрессин и окситоцин - первые гипофизарные гормоны с полностью установленной аминокислотной последовательностью. По своей химической структуре они представляют собой нонапептиды с одним дисульфидным мостиком.
Рассматриваемые гормоны дают разнообразные биологические эффекты: стимулируют транспорт воды и солей через мембраны, оказывают вазопрессорное действие, усиливают сокращения гладкой мускулатуры матки при родах, повышают секрецию молочных желез. Следует отметить, что вазопрессин обладает более высокой, чем окситоцин, антидиуретической активностью, тогда как последний сильнее действует на матку и молочную железу. Основным регулятором секреции вазопрессина является потребление воды, в почечных канальцах он связывается с рецепторами в цитоплазматических мембранах с последующей активацией в них фермента аденилатциклазы. За связывание гормона с рецептором и за биологический эффект отвечают разные участки молекулы.
Гипофиз, связанный через гипоталамус со всей нервной системой, объединяет в функциональное целое эндокринную систему, участвующую в обеспечении постоянства внутренней среды организма (гомеостаз). Внутри эндокринной системы гомеостатическая регуляция осуществляется на основе принципа обратной связи между передней долей гипофиза и железами-«мишенями» (щитовидная железа, кора надпочечников, гонады). Избыток гормона, вырабатываемого железой-«мишенью», тормозит, а его недостаток стимулирует секрецию и выделение соответствующего тропного гормона. В систему обратной связи включается гипоталамус. Именно в нем находятся чувствительные к гормонам желез-«мишеней» рецепторные зоны. Специфически связываясь с циркулирующими в крови гормонами и меняя ответную реакцию в зависимости от концентрации гормонов, рецепторы гипоталамуса передают свой эффект в соответствующие гипоталамические центры, которые координируют работу аденогипофиза, выделяя гипоталамические аденогипофизотропные гормоны. Таким образом, гипоталамус следует рассматривать как нейро-эндокринный мозг.